УДК 551.586
Сравнение фенологических моделей
определения начала пыления березы для
численного
прогнозирования переноса аллергенов
С.В.
Емелина, Е.В. Набокова, К.Г. Рубинштейн
Гидрометеорологический научно-исследовательский
центр
Российской Федерации, г. Москва, Россия
Рассмотрены возможности использования
фенологических моделей для определения даты начала пыления пыльцы березы на
Европейской территории России. В исследовании использованы температурно-временная,
последовательная, параллельная и переменная фенологические модели. Данные
моделирования сопоставлены с данными пыльцевого мониторинга в Санкт-Петербурге
(2012–2013 гг.) и Нижнем Новгороде (2009–2010 гг.). Показано, что в
рассматриваемых городах наиболее точно даты начала пыления определяет температурно-временная фенологическая модель.
Полученные результаты будут использованы в рамках задачи численного прогноза
переноса пыльцы березы.
Ключевые слова: фенологическая
модель, пыльца
березы, эмиссия пыльцы, прогноз начала пыления
березы
Comparison of phenological models
for determining the beginning of birch pollen dispersion
for numerical forecasting of allergens transport
S.V. Emelina, E.V.
Nabokovа, K.G. Rubinstein
Hydrometeorological Research
Center of Russian Federation, Moscow, Russia
The possibilities of using phenological models to determine the date of
the beginning of birch pollen dispersion in the European part of Russia are
considered. The study uses the temperature-time, sequential, parallel, and
alternating phenological models. Simulation data are compared
with pollen monitoring data for St. Petersburg (2012–2013) and Nizhny Novgorod
(2009–2010). It is shown that the temperature-time
phenological model is the most accurate in determining the dates of the
start of pollen dispersion in the cities. The results can be useful for the
numerical prediction of birch pollen transport.
Keywords: phenological model, birch pollen, pollen
emission, birch pollen dispersion forecasting
Введение
Зерна пыльцы различных растений, попадая в организм человека, могут
вызывать аллергические реакции – поллиноз. По данным Всемирной организации
здравоохранения, около 15 % населения Европы страдает
от поллиноза [25]. В весенний период основной причиной поллиноза на территории
Европы является пыльца березы. По данным [10],
в Центральной и Западной Европе чувствительность к пыльце березы наблюдается у
37,6 % людей, страдающих поллинозом. Березовые леса имеют обширную географию
распространения. В Европе береза
встречается
от Скандинавии [24] до центральной Испании, на севере Греции и на юге Италии
[21], в Хорватии [15, 23], а также в западной и центральной России [1, 21].
Самые высокие среднегодовые концентрации пыльцы березы фиксируются в Финляндии,
Литве, Латвии, Эстонии, Польше, России
и Белоруссии [18].
Даты начала пыления березы в различных регионах заметно варьируются.
Так, на севере Испании оно начинается в феврале и марте, в
западной Европе – в конце марта. В центральной Европе средние даты начала пыления
приходятся на первую половину апреля, в Северной
Европе цветение березы продолжается с конца апреля по май [6, 8, 14,
16, 17].
Отличительной характеристикой пыльцы березы
является возможность переноса на расстояния до 1000 км [22], что в сочетании
с обширным распределением этого вида и вариативностью даты начала пыления
делает ее наиболее опасным аллергеном. Таким образом, широкое распространение
главной причины поллиноза – березы, а также растущее число случаев поллиноза в
России [2] свидетельствуют
о высокой актуальности своевременного прогноза начала пыления и переноса пыльцы
аллергенов в течение всего вегетационного периода.
Основным инструментом для анализа
и прогноза концентрации пыльцы в воздухе является численное моделирование,
которое объединяет в себе фенологическую и транспортную модели. В данной работе
приведено описание различных типов фенологических моделей, которые используются
для прогноза начала и конца пыления, а также сопоставление прогностических дат
начала пыления березы с фактическими данными пыльцевого мониторинга на
Европейской территории России.
Типы фенологических моделей
Для определения начала пыления используются
фенологические модели, которые описывают стадии развития растений в
зависимости от изменений метеорологических параметров. Для их типизации
необходимо рассмотреть два температурных периода, влияющих на развитие растений
– период охлаждения и период температурного форсинга.
Охлаждение (яровизация) – это период воздействия низких положительных
или слабоотрицательных температур на растение поздней осенью. Яровизация
растений вызвана их адаптацией к сезонным температурным изменениям.
Температура, при которой происходит яровизация, зависит от вида и сорта
растений. В средних широтах она происходит при температурах от -3,5 °С
до 10,2 °С и достигает оптимума при температуре +3,5 °С.
Температурный форсинг – это период накопления тепла,
необходимого для раскрытия почек. Процессы температурного форсинга и яровизации
математически описываются с помощью соответствующих функций и представляют
собой суммированные по времени уровни форсинга ![]() и охлаждения
и охлаждения ![]() , рассчитывающиеся разными способами в зависимости
от среднесуточной температуры воздуха:
, рассчитывающиеся разными способами в зависимости
от среднесуточной температуры воздуха:
![]() ,
(1)
,
(1)
где ![]() – функция
температурного форсинга;
– функция
температурного форсинга; ![]() – уровень форсинга;
– уровень форсинга; ![]() –
начальная дата периода форсинга (накопления тепла);
–
начальная дата периода форсинга (накопления тепла); ![]() – конечная дата периода форсинга.
– конечная дата периода форсинга.
![]() ,
(2)
,
(2)
где ![]() – функция
охлаждения;
– функция
охлаждения; ![]() – уровень охлаждения;
– уровень охлаждения; ![]() – начальная дата периода охлаждения;
– начальная дата периода охлаждения; ![]() – конечная дата периода охлаждения.
– конечная дата периода охлаждения.
В зависимости от описания периодов охлаждения и форсинга фенологические
модели можно разделить на четыре группы [7].
1. Температурно-временная модель,
или однофазная модель [3, 13].
Данная модель не учитывает период охлаждения, рассматривается только период
накопления тепла, который начинается от фиксированной даты. Уровень форсинга ![]() принимает нулевое значение при
среднесуточной температуре ниже базовой и равен разности между среднесуточной и
базовой температурами, когда среднесуточная температура превышает базовую:
принимает нулевое значение при
среднесуточной температуре ниже базовой и равен разности между среднесуточной и
базовой температурами, когда среднесуточная температура превышает базовую:
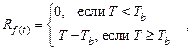 (3)
(3)
где ![]() – базовая
температура.
– базовая
температура.
Когда функция форсинга достигает порогового значения, начинается выброс
пыльцы. Для умеренных широт значение базовой температуры составляет 5 °С
согласно [3] и 3,5 °С согласно [13], а пороговое значение функции форсинга
– 70 °С.
2. Последовательная модель. В
последовательной модели [11–19] скорость охлаждения описывается треугольной
функцией принадлеж-ности и определяется тремя значениями
температуры периода охлаждения – минимальной, оптимальной и максимальной:
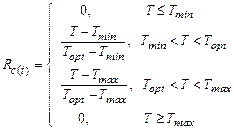 ,
(4)
,
(4)
где Tmin, Topt, Tmax – минимальная,
оптимальная и максимальная температура, °С.
Оптимальная температура для уровня охлаждения, как и базовая температура
для определения уровня накопления тепла, при этом определяется для каждого
конкретного вида отдельно. Уровень форсинга ![]() описывается функцией с тремя
оптимизируемыми параметрами (a, b, c):
описывается функцией с тремя
оптимизируемыми параметрами (a, b, c):
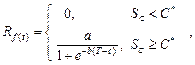 (5)
(5)
где a, b, c – константы, определяемые для каждого вида растения; ![]() – температурный порог охлаждения, °С.
– температурный порог охлаждения, °С.
3. В параллельной модели
предполагается, что процесс форсинга может идти во время периода охлаждения [9, 12]. Уровень охлаждения рассчитывается аналогично последовательной модели по
формуле (4). Уровень форсинга рассчитывается по формуле (6). С помощью
дополнительных коэффициентов ![]() и
и ![]() уровень охлаждения влияет на интенсивность
накопления тепла.
уровень охлаждения влияет на интенсивность
накопления тепла.
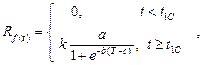 (6)
(6)
где a, b, c – константы, определяемые для каждого вида растения; ![]() – дополнительный коэффициент, который
определяется как
– дополнительный коэффициент, который
определяется как
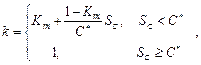 (7)
(7)
где ![]() – коэффициент,
определяемый для каждого вида растения;
– коэффициент,
определяемый для каждого вида растения; ![]() –
температурный порог охлаждения, °С.
–
температурный порог охлаждения, °С.
4. Переменная модель является
усложненной температурно-временной моделью [5]. С 1 января
начинается накопление уровня
форсинга, который рассчитывается по формуле (3), однако пороговое значение для
функции форсинга не является константой, а имеет обратную экспоненциальную
зависимость от функции охлаждения:
![]() ,
(8)
,
(8)
где ![]() – начальная дата
периода форсинга (накопления тепла);
– начальная дата
периода форсинга (накопления тепла); ![]() – конечная дата периода форсинга;
– конечная дата периода форсинга; ![]() – уровень охлаждения; a, b, c – константы, определяемые для каждого вида
растения.
– уровень охлаждения; a, b, c – константы, определяемые для каждого вида
растения.
При использовании представленных выше моделей
необходим подбор параметров для конкретного географического региона на основе
данных наблюдений. В [7] приведены значения для
параметров последовательной модели для Франции и Бельгии. Они представлены в
табл. 1. В табл. 2
представлены страны разработки фенологической модели, периоды наблюдений и число станций, по
которым были выведены необходимые для расчета константы.
Таблица 1. Значения
параметров и коэффициентов последовательной фенологической модели
Table 1. Values of parameters and coefficients of
sequential phenological model
|
Страна |
Tmin |
Topt |
Tmax |
C* |
a |
b |
c |
|
Франция
|
-3,4 |
|
10,4 |
|
28,4 |
0,185 |
18,4 |
|
Бельгия
|
-10,8 |
8,6 |
22,9 |
403,6 |
66,1 |
0,35 |
7,88 |
Примечание. Tmin,
Topt,
Tmax – минимальная, оптимальная и максимальная температуры; C* – температурный порог охлаждения; a,
b, c – константы, определяемые для каждого вида растения.
Таблица 2. Период наблюдений и число станций, на основе
наблюдений
на которых разрабатывались фенологические модели
Table
2. The observation period and the
number of stations using for phenological models developing
|
Фенологическая модель |
Страна |
Данные измерений |
|
|
Число станций |
Период измерений |
||
|
ТТМ |
Великобритания |
14 |
1960–1980 |
|
SM (F) |
Франция |
1 |
1974–1992 |
|
SM (B) |
Бельгия |
1 |
сезон 2009–2010 |
|
PM |
Франция |
1 |
1974–1992 |
|
AM |
Бельгия |
1 |
сезон 2009–2010 |
Примечание. TTM – температурно-временная
модель; SM –
последовательная модель; PM – параллельная модель, AM – переменная модель.
Данные пыльцевого мониторинга
Результаты моделирования
сравнивались с данными пыльцевого
мониторинга, проводимого Российской Ассоциацией Аллергологов и Клинических
иммунологов (РААКИ), МГУ и фармацевтической компанией «Никомед». Архив
ежедневных данных в текстовом формате по концентрации
аллергенов (кол-во ядер/м3) находился ранее в открытом доступе для
Санкт-Петербурга (доступный период наблюдений 2012–2013 гг.) и для Нижнего Новгорода
(период наблюдений 2009–2010 гг.). На данных рядах фактических наблюдений были проведены оценки качества
воспроизведения даты начала пыления березы по различным типам фенологических
моделей.
Сравнительная оценка
фенологических моделей
Для оценки точности
фенологических моделей были рассчитаны даты начала пыления березы в Нижнем
Новгороде и Санкт-Петербурге. В качестве необходимых метеорологических
параметров были использованы срочные данные с соответствующих синоптических
станций дискретностью 3 часа: Нижний Новгород (56,3° с. ш.,
44,0° в. д.) и Воейково (60,0° с. ш.,
30,3° в. д.). Были рассмотрены четыре типа моделей: температурно-временная
(с базовой температурой в +5 °С), последовательная модель в двух вариантах (с параметрами
форсинга для Франции и Бельгии), параллельная
модель и переменная. Появление первых ядер пыльцы в атмосферном воздухе
рассматривалось как начало пыления. Результаты представлены в табл. 3 и 4. В
скобках указано число дней отставания/опережения прогностической даты начала
пыления от фактической.
Согласно представленным
данным можно сделать вывод, что в Нижнем Новгороде наиболее точно дату начала
пыления описывает температурно-временная модель. За два рассматриваемых года
дата начала запаздывает от фактической на 3–5 дней. Более поздние даты начала
пыления (более чем на 10 дней) получаются при использовании параллельной, переменной
и последовательной модели (Франция). Последовательная модель (Бельгия), наоборот,
воспроизводит даты начала пыления березы с «запасом» в 13–14 дней.
Похожие результаты с
опережением фактической даты наблюдаются в Санкт-Петербурге: в 2013 г. на
29 дней раньше, в 2012 г. – на 12 дней. При этом, при использовании
параллельной модели береза в Санкт-Петербурге в рассматриваемые годы не
начинает пылить (сумма накопленных температура не достигает необходимого
порога), а по переменной модели начало пыления в 2012/2013 гг наступает на
54/37 дней позже. Несогласованный результат получается с температурно-временной
моделью: в 2012 г. она опережает фактические значения на 14 дней, в 2013
году – запаздывает на 11 дней.
Таблица 3. Фактические и расчетные даты начала пыления березы
в Нижнем Новгороде, 2009-2010 гг.
Table 3. Actual
and estimated dates of the start of dusting of birch in Nizhny Novgorod,
2009-2010
|
Год |
Факт |
ТТМ |
SM (F) |
SM (B) |
PM |
AM |
|
2009 |
29.04 |
03.05 (+5) |
07.05 (+9) |
15.04 (-14) |
14.05 (+15) |
06.06 (+38) |
|
2010 |
19.04 |
22.04 (+3) |
03.05 (+14) |
06.04 (-13) |
10.05 (+21) |
02.06 (+44) |
Примечание. Факт –
фактические наблюдения; TTM – температурно-временная
модель; SM –
последовательная модель (F –Франция, B – Бельгия); PM – параллельная модель; AM – переменная модель.
Таблица 4. Фактические и расчетные даты начала пыления березы
в Санкт-Петербурге, 2012-2013 гг.
Table 4. Actual and estimated dates of the start of dusting of birch
in St.Petersburg, 2012-2013
|
Год |
Факт |
ТТМ |
SM (F) |
SM (B) |
PM |
AM |
|
2012 |
22.04 |
06.05 (+14) |
15.05 (+23) |
10.04 (-12) |
– |
15.06 (+54) |
|
2013 |
17.05 |
08.05 (-11) |
25.04 (-20) |
06.04 (-29) |
– |
23.06 (+37) |
Примечание. Факт –
фактические наблюдения; TTM – температурно-временная
модель; SM –
последовательная модель (F –Франция, B – Бельгия); PM – параллельная модель; AM – переменная модель.
Несовпадение результатов расчетных и фактических
дат, во-первых, может быть обусловлено тем, что по фактическим данным появление
первых ядер пыльцы березы в атмосферном воздухе может являться не началом
пыления в рассматриваемом пункте, а фактом заноса из более южных широт. Во-вторых,
используемые эмпирические показатели были разработаны для конкретных областей и
широт, что, вполне возможно, ограничивает использование данных формул в других
районах. Сильные различия в фактических датах начала пыления в Санкт-Петербурге
с 2012 и 2013 гг. обусловлены особенностями температурного режима. Так, сезон
цветения березы в 2013 г. в Европе начался на 2–3 недели позже, чем
обычно, из-за поздней весны. Учитывая данные особенности, можно сказать, что
точнее всего даты начала пыления в рассматриваемых городах описывает
температурно-временная модель.
Список литературы
1. Минаева Н.В., Новоселова Л.В., Плахина К.А.,
Ширяева Д.М. Пыльцевая сенсибилизация и аэропалинологический
мониторинг в определении значимых аллергенов при раннем весеннем поллинозе //
Российский аллергологический журнал. 2015. № 2. С. 19-24.
2. Ширяева
Д.М., Минаева Н.В., Новоселова Л.В. Экологические
аспекты поллинозов. Обзор литературы // Экология человека. 2016. № 12. С.
3-10.
3. Cannell M.G.R., Smith R.I. Thermal Time, Chill Days and Prediction of Budburst
in Picea-Sitchensis // Journal of Applied Ecology. 1983. No. 20. P. 951-963.
4. Charalampopoulos A., Damialis A.,
Tsiripidis I., Mavrommatis T., Halley J.M., Vokou D. Pollen
production and circulation patterns along an elevation gradient in Mt Olympos
(Greece) National Park // Aerobiologia. 2013. Vol. 29 (4). P. 455-472.
5. Chuine
I. A unified model for
budburst of trees // J. Theoretical Biology. 2000. Vol. 207. P. 337-347.
6. Emberlin J., Detandt M., Gehrig R., Jaeger
S., Nolard N., Rantio-Lehtimäki A. Re-
sponses in the start of Betula (birch) pollen seasons to recent changes in
spring temperatures across Europe // Int. J. Biometeorol. 2002. Vol. 46.
P. 159-170.
7. Fu Y.H.,
Campioli M., Deckmyn G., Janssens I.A.
The Impact of Winter and Spring Temperatures on
Temperate Tree Budburst Dates: Results from an Experimental Climate Manipulation
// PLoS ONE. 2012. Vol. 7, no. 10. P. e47324.
8. Grewling Ł., Jackowiak B., Nowak M., Uruska A., Smith M.
2012. Variations and trends of birch pollen seasons during 15 years (1996–2010)
in relation to weather conditions in Poznań(Western Poland) // Grana.
Vol. 51, no. (4). P. 280-292.
9. Hanninen H. Modelling bud dormancy release in trees from cool and
temperate regions // Acta for Fenn. 1990. Vol. 213. P. 1-47.
10. Heinzerling L.M., Burbach G.J.,
Edenharter G. et al. GALEN skin test study I: GALEN
harmonization of skin prick testing: novel sensitization patterns for inhalant
allergens in Europe // Allergy. 2009. Vol. 64, no. 10. P. 1498-1506.
11. Kramer
K. Selecting a Model to
Predict the Onset of Growth of Fagus-Sylvatica // J. Applied Ecology. 1994.
Vol. 31. P. 172-181.
12. Landsberg
J.J. Apple Fruit Bud
Development and Growth – Analysis and an Empirical Model // Annals of Botany.
1974. Vol. 38. P. 1013-1023.
13. Linkosalo
T., Ranta H., Oksanen A., Siljamo P., Luomajoki A., Kukkonen J., Sofiev M. A double-threshold temperature sum model for
predicting the flowering duration and relative intensity of Betula pendula and
B. pubescens // Agr. Forest Meteorol. Vol. 150. P. 1579-1584.
doi:10.1016/j.agrformet.2010.08.007,2010.
14. Melgar
M., Trigo M.M., Recio M., Docampo S., Gracía-Sánchez J., Cabezudo
B. Atmospheric pollen dynamics in
Münster, north-western Germany: a three-year study (2004–2006) //
Aerobiologia. 2012. Vol. 28. P. 423-434.
15.
Peternel R., Milanović
S.M., Hrga I., Mileta T.,Čulig J. Incidence
of Betulaceae pol-len and pollinosis in Zagreb, Croatia, 2002–2005 // Ann.
Agric. Environ. Med. 2007. Vol. 14. P. 87-91.
16. Piotrowska
K. Pollen production in selected species of anemophilous
plants // Acta Agrobot. 2008. Vol. 61, no. 1. P. 41-52.
17. Piotrowska-Weryszko
K., Weryszko-Chmielewska E. The airborne pollen calendar for
Lublin, central-eastern Poland. Ann. Agric // Environ. Med. 2014. Vol. 21, no.
3. P. 541-545.
18. Puc M., Wolski T.,
Câmara Camacho I., Myszkowska D. et al. Fluctuation of birch
(Betula L.) pollen seasons in Poland // Acta Agrobot. 2015. Vol. 68, no. 4. P.
303-313.
19.
Sarvas
R. Investigations on the
annual cycle of development of forest trees. II. Autumn dormancy and winter
dormancy // Commun Inst For Fenn. 1974. Vol. 84. P. 1-101.
20. Ščevková
J., Dušička J., Chrenová J. Annual pollen
spectrum variations in the air of Bratislava (Slovakia): years 2002–2009
// Aerobiologia. 2010. Vol. 26. P. 277-287.
21.
Skjøth C.A.,
Šikoparija B., Jäger S. EAN-Network, Pollen sources //
Allergenic pollen: a review of the production, release, distri- bution and
health impacts. Springer: London, 2013. P. 9-27.
22. Sofiev M., Siljamo P., Ranta
H. Towards numerical forecasting of long-range air transport of birch pollen:
theoretical considerations and a feasibility study // Int. J. Biometeorol.
2006. Vol. 50. P. 392.
23.
Stefanic E., Rasic S., Merdic
S., Colakovic K. Annual variation of airborne pollen in the city
of Vinkovci, northeastern Croatia // Ann. Agric. Environ. Med. 2007. Vol. 14.
P. 97-101.
24. Yli-Panula
E., Fekedulegn D.B., Green B.J., Ranta H. Analysis of
airborne Betula pollen in Finland; a 31-year perspective // Int. J. Environ.
Res. Public Health. 2009. Vol. 6. P. 1706-1723.
25. Huynen
M., Menne B., Behrendt H., Bertollini R. et al. Phenology and human health: allergic disorders //
Health and Global Environmental Change Series. 2003. No. 1. EUR/o3/5036791.
55 p.
References
1. Minaeva N.V., Novoselova L.V., Plakhina K.V.,
Shiryaeva D.M. Pollen sensitization and aeropalynological
monitoring for determination of significant allergens in early spring
pollinosis. Rossiiskii allergologicheskii zhurnal [Russian Allergology Journal], 2015, no. 2, pp. 19-24.
[in Russ].
2. Shiriaeva D.M., Minaeva N.V., Novoselova L.V. Ecological aspects of pollinosis. Literature review. Ekologiya cheloveka [Human Ecology], 2016, no. 12, pp. 3-10. [in
Russ].
3. Cannell M.G.R., Smith R.I.
Thermal Time, Chill Days and Prediction of Budburst in Picea-Sitchensis. Journal of Applied Ecology, 1983, no.
20, pp. 951-963.
4. Charalampopoulos
A., Damialis A., Tsiripidis I., Mavrommatis T., Halley J.M., Vokou D. Pollen production
and circulation patterns along an elevation gradient in Mt Olympos (Greece)
National Park. Aerobiologia, 2013,
vol. 29, no. 4, pp. 455-472.
5. Chuine I. A unified model
for budburst of trees. J. Theoretical
Biology, 2000, vol. 207, pp. 337-347.
6. Emberlin J.,
Detandt M., Gehrig R., Jaeger S., Nolard N., Rantio-Lehtimäki A. Re-sponses in the start of
Betula (birch) pollen seasons to recent changes in spring temperatures across
Europe. Int. J. Biometeorol., 2002, vol. 46, pp. 159-170.
7. Fu Y.H.,
Campioli M., Deckmyn G., Janssens I.A.
The Impact of Winter and Spring Temperatures on Temperate Tree Budburst Dates:
Results from an Experimental Climate Manipulation. PLoS ONE, 2012, vol. 7, no. 10, p. e47324.
8. Grewling
Ł., Jackowiak B., Nowak M., Uruska A., Smith M. 2012. Variations
and trends of birch pollen seasons during 15 years (1996–2010) in relation to
weather conditions in Poznań(Western Poland). Grana, vol. 51, no. 4, pp. 280-292.
9. Hanninen H.
Modelling bud dormancy release in trees from cool and temperate regions. Acta for Fenn., 1990, Vol. 213, pp.
1-47.
10. Heinzerling L.M., Burbach G.J., Edenharter G. et al. GALEN skin test
study I: GALEN harmonization of skin prick testing: novel sensitization
patterns for inhalant allergens in Europe. Allergy,
2009, vol. 64, no. 10, pp. 1498-1506.
11.
Kramer K. Selecting a Model to Predict the Onset of Growth of Fagus-Sylvatica. J.
Applied Ecology., 1994, vol. 31, pp. 172-181.
12.
Landsberg J.J. Apple Fruit Bud Development and Growth – Analysis and an Empirical
Model. Annals of Botany, 1974, vol.
38, pp. 1013-1023.
13.
Linkosalo T.,
Ranta H., Oksanen A., Siljamo P., Luomajoki A., Kukkonen J., Sofiev M. A double-threshold temperature sum model for predicting the flowering
duration and relative intensity of Betula pendula and B. pubescens. Agr. Forest Meteorol., vol. 150, pp.
1579-1584. Doi:10.1016/j.agrformet.2010.08.007,2010.
14. Melgar M., Trigo M.M., Recio M., Docampo S.,
Gracía-Sánchez J., Cabezudo B. Atmospheric pollen dynamics in
Münster, north-western Germany: a three-year study (2004–2006). Aerobiologia, 2012, vol. 28, pp. 423-434.
15. Peternel R., Milanović S.M., Hrga I., Mileta
T.,Čulig J. Incidence
of Betulaceae pol-len and pollinosis in Zagreb, Croatia, 2002–2005. Ann. Agric. Environ. Med., 2007, vol. 14, pp. 87-91.
16. Piotrowska K. Pollen production in selected
species of anemophilous plants. Acta
Agrobot, 2008, vol. 61, no. 1, pp. 41-52.
17. Piotrowska-Weryszko K., Weryszko-Chmielewska E. The airborne
pollen calendar for Lublin, central-eastern Poland. Ann. Agric. Environ. Med., 2014, vol. 21, no. 3, pp.
541-545.
18. Puc M., Wolski T., Câmara Camacho I., Myszkowska D. et al. Fluctuation of birch (Betula L.) pollen
seasons in Poland. Acta Agrobot,
2015, vol. 68, no. 4, pp. 303-313.
19. Sarvas R. Investigations on the annual cycle of development of forest trees. II.
Autumn dormancy and winter dormancy. Commun
Inst For Fenn., 1974, vol. 84, pp. 1-101.
20. Ščevková J., Dušička J.,
Chrenová J. Annual pollen spectrum variations in the air of Bratislava
(Slovakia): years 2002–2009. Aerobiologia,
2010, vol. 26, pp. 277-287.
21. Skjøth C.A., Šikoparija B., Jäger S. EAN-Network, Pollen
sources. Allergenic pollen: a review of the production, release, distri- bution
and health impacts. Springer: London, 2013, pp. 9-27.
22.
Sofiev M., Siljamo P., Ranta
H. Towards numerical forecasting of
long-range air transport of birch pollen: theoretical considerations and a
feasibility study. Int. J. Biometeorol.,
2006, vol. 50, pp. 392.
23. Stefanic E., Rasic S., Merdic S., Colakovic K. Annual variation of
airborne pollen in the city of Vinkovci, northeastern Croatia. Ann. Agric. Environ. Med., 2007, vol.
14, pp. 97-101.
24. Yli-Panula E., Fekedulegn D.B., Green B.J., Ranta H. Analysis of
airborne Betula pollen in Finland; a 31-year perspective. Int. J. Environ. Res. Public Health., 2009, vol. 6, pp. 1706-1723.
25.
Huynen M., Menne
B., Behrendt H., Bertollini R. et al.
Phenology and human health: allergic disorders. Health and Global Environmental Change Series, 2003, no. 1,
EUR/o3/5036791, 55 p.
Поступила в редакцию 05.06.2019 г.
Received by the editor 05.06.2019.